Ионные каналы (ИК) клеточной мембраны имеют огромное значение для жизни клеток. Они обеспечивают обмен клетки с окружающей средой веществом, энергией и информацией, с них начинаются и ими поддерживаются процессы возбуждения и торможения в нервной системе и мышцах, именно они (вместе и другими молекулярными рецепторами) обеспечивают восприятие клеткой внешних сигналов. С помощью ИК происходит передача в клетку управляющих сигналов из окружающей её среды. Именно ИК обеспечивают синаптическую передачу возбуждения от возбуждённого нейрона на другие клетки. Обобщая, можно сказать, что почти все важнейшие физиологические процессы в организме начинаются с ионных каналов и поддерживаются ими!
Итак, ионные каналы (ИК) - это сложные трансмембранные белковые структуры, пронизывающие клеточную мембрану поперёк в виде нескольких петель и образующие в мембране сквозное отверстие (пору). Канальные белки состоят из субъединиц, образующих структуру со сложной пространственной конфигурацией, в которой кроме поры обычно имеются дополнительные молекулярные системы: открытия, закрытия, избирательности, инактивации, рецепции и регуляции. ИК могут иметь не один, а несколько участков (сайтов) для связывания с управляющими веществами (лигандами).
Ионные каналы можно рассматривать как транспортный механизм, обеспечивающий перемещение ионов между цитоплазмной клетки и наружной средой.
Кроме ИК в мембране суцществуют и другие транспортные системы для переноса через неё различных веществ (смотрите транспортные механизмы мембраны). Так, перенос веществ может осуществляться специальными транспортными белками, или транслоказами. Транслоказы - это несколько иное понятие, чем ИК. В отличие от мембранных каналов, транслоказы в процессе переноса вещества через мембрану взаимодействуют с ним как с лигандом и при этом претерпевают конформационные изменения. По кинетике перенос веществ с помощью транслоказ в виде облегчённой диффузии напоминает ферментативную реакцию.
Упрощённое определение:
Ионные каналы - это поры (дырочки) в клеточной липидной мембране, которые "обшиты" по краям белковой нитью, чтобы дырочки не затянулись. Эти поры могут становиться пошире или поуже: либо сами по себе, либо при определённых воздействиях. Каналы могут иметь разное строение, поэтому разные виды каналов имеют разную проницаемость, избирательность и управляемость.
Итак, ионный канал - это интегральный белок, образующий в мембране пору для обмена клетки с окружающей средой ионами K+, Na+, H+, Ca2+, Cl-, а также водой, и способный изменять свою проницаемость.
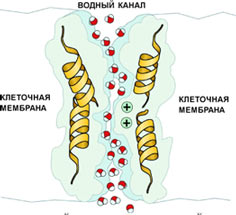
Аквапорины - водные неионные каналы мембраны
В мембране существуют и неионные каналы. Например, аквапорины — это специальные водные каналы, пропускающие через себя воду. Это тоже мембранные каналы, хотя их формально нельзя назвать "ионными каналами".
Пространственная структура канального белка-аквапорина представляет собой цилиндрический канал, по которому движутся молекулы воды. Через него проходит вода и только вода, но не ионы. Аминокислоты в этом белке расположены таким образом, что полярность создаваемого ими электростатического поля переключается в центре канала на обратную. Поэтому молекулы воды, дойдя до середины канала, переворачиваются так, что их дипольные моменты в верхней и нижней части канала оказываются направленными в противоположные стороны. Такое переориентирование предотвращает просачивание через канал заряженных ионов. Аквапорин не пропускает даже ионы гидроксония H3O+ (то есть гидратированные протоны, или ионы водорода), от концентрации которых зависит кислотность среды. При этом клеточный мембранный "водопровод" обладает потрясающей пропускной способностью: он пропускает до миллиарда молекул воды в секунду. Сейчас известно уже около 200 разновидностей белков водных каналов у растений и животных, в том числе 11 - у человека. Благодаря аквапоринам клетки не только регулируют свой объём и внутреннее давление, но и выполняют такие важные функции, как всасывание воды в почках животных и корешках растений.
В настоящее время в молекулярной биологии в основном завершён описательный период в исследовании многообразия катион-транспортирующих ионных каналов в клетках эукариот. Теперь на первый план выходят проблемы познания механизмов регуляции ионных каналов и описание их участия в реакциях живой клетки на различные воздействия и на изменение её микроокружения.
Регулирумый перенос ионов через гидрофильные поры мембраны с помощью управляемых ИК является важнейшим свойством живых клеток, как электровозбудимых, так и невозбудимых.
В связи с этим целесообразно использовать в классификации ионных каналов именно принцип управления их деятельностью. Принцип управления состоянием ионных каналов и был положен в основу предложенной нами (Сазонов В.Ф., 2011.) функциональной классификации ионных каналов.
Видео: Ионные каналы в мембране
Строение ИК
ИК состоят из белков сложной структуры (белков-каналоформеров). Схематические изображения ИК приведены ниже, например: натриевый НАХ-рецепторный ионный канал.
На рисунке справа показан натриевый канал: вид сверху, с наружной стороны мембраны (Источник: Horn R. (2011). Peering into the spark of life. Nature475, 305–306).
Белки ИК имеют определённую конформацию, образующую трансмембранную пору, и "вшиты" в липидный слой мембраны. Канальный белковый комплекс может состоять либо из одной белковой молекулы, либо из нескольких белковых субъединиц, одинаковых или разных по строению. Эти субъединицы могут кодироваться разными генами, синтезироваться на рибосомах по-отдельности и затем собираться в виде целостного канала. В другом случае канал может представлять собой единый полипептид, который в виде петель прошивает мембрану несколько раз. На начало XXI века известно более 400 белков-каналоформеров, для биосинтеза которых используется 1-2% генома человека.
Домены - это отдельные компактно оформленные части канального белка или субъединиц. Сегменты - это части белкка-каналоформера, свёрнутые спирально и прошивающие мембрану. Концевые домены белка-каналоформера (N- и С-терминальные домены) могут торчать из мембраны как наружу, так и внутрь клетки.
Практически все ИК имеют в составе своих субъединиц регуляторные домены, способные связываться с различными управляющими веществами (регуляторными молекулами) и за счёт этого менять состояние или свойства канала. В потенциал-активируемых ИК один из трансмембранных сегментов содержит специальный набор аминокислот с положительными зарядами и работает как сенсор электрического потенциала мембраны. При изменении потенциала такой сенсор меняет состояние канала с открытого на закрытое или наоборот. Таким образом, ИК могут управляться определёнными воздействиями извне, это важное их свойство.
ИК в своём составе могут иметь также вспомогательные субъединицы, выполняющие модуляторные, структурные или стабилизирующие функции. Один класс таких субъединиц - внутриклеточные, расположенные полностью в цитоплазме, а второй - мембранные, т.к. они имеют трансмембранные домены, прошивающие мембрану.
По структуре ИК возможно провести их классификацию, о чём будет сказано ниже.
Свойства ИК
Селективность - это избирательная повышенная проницаемость ИК для определённых ионов. Для других ионов проницаемость понижена. Такая избирательность определяется селективным фильтром - самым узким местом канальной поры. Фильтр, кроме узких размеров, может иметь также локальный электрический заряд. Например, катион-селективные каналы обычно имеют в области своего селективного фильтра отрицательно заряженные остатки аминокислот в составе белковой молекулы, которые притягивают положительные катионы и отталкивают отрицательные анионы, не пропуская их через пору.
Управляемая проницаемость - это способность ИК открываться или закрываться при определённых управляющих воздействиях на канал. Понятно, что закрытый канал имеет пониженную проницаемость, а открытый - повышенную. По этому свойству ИК можно классифицировать в зависимости от способов их открытия: например, потенциал-активируемые, лиганд-активируемые и т.д.
Инактивация - это способность ИК через некоторое время после своего открытия автоматически понижать свою проницаемость даже в том случае, когда открывший их активирующий фактор продолжает действовать.
Быстрая инактивация - это особый процесс со своим особым механизмом, отличающийся от медленного закрытия канала (медленной инактивации). Закрытие (медленная инактивация) канала происходит за счёт процессов, противоположных процессам, обеспечившим его открытие, т.е. за счёт изменения конформации канального белка. А вот, например, у потенциал-активируемых каналов быстрая инактивация происходит с помощью специальной молекулярной "пробки-затычки", напоминающей пробку на цепочке, которую обычно используют в ваннах. Эта пробка представляет собой аминокислотную (полипептидную) петлю с утолщением на конце в виде трёх аминокислот, которым и затыкается внутреннее устье канала со стороны цитоплазмы. Именно поэтому потенциал-зависимые ИК для натрия, обеспечивающие развитие потенциала действия и движение нервного импульса, могут пропускать в клетку ионы натрия только в течение нескольких миллисекунд, а затем они автоматически закрываются своими молекулярными пробками, несмотря на то, что открывающая их деполяризация продолжает действовать. Другим механизмом инактивации ИК может служить модификация дополнительными субъединицами внутриклеточного устья канала.
Блокировка - это способность ИК под действием веществ-блокаторов фиксировать какое-то одно своё состояние и не реагировать на обычные управляющие воздействия. В таком состоянии канал просто перестаёт давать ответы на управляющие воздействия. Блокировку вызывают вещества-блокаторы, которые могут называться антагонистами, блокаторами или литиками.
Антагонисты - это вещества, препятствующие активирующему действию других веществ на ИК. Такие вещества способны хорошо связываться с рецепторным участком ИК, но не способны изменить состояние канала, вызвать его ответную реакцию. Получается блокада рецептора и вместе с ним - блокада ИК. Следует помнить, что антагонисты не обязательно вызывают полную блокаду рецептора и его ИК, они могут действовать более слабо и лишь ингибировать (угнетать) работу канала, но не прекращать её полностью
Агонисты-антагонисты - это вещества, которые обладают слабым стимулирующим влиянием на рецептор, но при этом блокируют действие естественных эндогенных управляющих веществ.
Блокаторы- это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином - адреноблокаторы; гистамина - гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
Литики - это те же блокаторы в отношении ИК, но этот термин более старый. Он используется как синоним для блокатора: холинолитик, адренолитик и т.д. В то же время в фармакологии термин "литик" применяется в более широком смысле - как вещество, препятствующее не только действию лиганда, а вообще препятствующее передаче возбуждения.
Пластичность - это способность ИК изменять свои свойства, свои характеристики. Наиболее распространённый механизм, обеспечивающий пластичность - это фосфорилирование аминокислот канальных белков с внутренней стороны мембраны ферментами-протеинкиназами. К канальным белкам присоединяются фосфорные остатки от АТФ или ГТФ - и канал меняет свои свойства. Накпример, фиксируется в постоянно закрытом состоянии, или, наоборот, в открытом.
Функции ИК
Главная функция ИК - обеспечивать управляемое перемещение ионов через мембрану.
В зависимости от проходящих через них ионов ИК подразделяют на натриевые, калиевые, кальциевые, хлорные, протонные (водородные).
Функции ИК:
1. Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление и защелачивание.
3. Регуляция ионного обмена (обмен солей): изменение внутриклеточного ионного состава и концентрации.
4. Создание и изменение мембранных потенциалов: потенциал покоя; в возбудимых клетках - локальные потенциалы, потенциал действия.
5. Проведение возбуждения в возбудимых клетках: обеспечение движения нервных импульсов.
6. Трансдукция в сенсорных рецепторах: преобразование раздражения (стимула) в возбуждение.
7. Управление активностью клетки: за счёт обеспечения потоков вторичного мессенджера - Са2+.
Функциональные состояния ИК
1. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал закрыт и ионы не проходят через него.
3. Активированное. Канал может выполнять свои функции, т.е. открываться и закрываться под действием его регуляторов (управляющих веществ или электрических потенциалов).
4. Инактивированное. Канал не может выполнять свои функции, т.е. открываться и закрываться, он "фиксируется" в каком-то одном состоянии.
5. Блокированное. Канал перекрыт, инактивирован веществом-антагонистом (блокатором), занявшем место управляющего вещества.
6. Модулированное (фосфорилированное). Канал изменяет свои обычные свойства под действием фосфорилирования - присоединения к какому-то его участку фосфатного остатка.
Структурно-функциональные нарушения ИК
Каналопатии - это группа врождённых заболеваний, вызванных нарушениями в работе ИК. Каждая каналопатия обусловлена патологией соответствующих ИК. В основе патологии лежит либо мутация генов, кодирующих строение белков-каналоформеров, либо производство аутоантител, агрессивных по отношению к собственным ИК.
Как уже говорилось выше, ИК можно классифицировать различным образом:
1. По селективности (степени избирательной проницаемости к определённым ионам). В этом случае мы будем говорить о натриевых, калиевых, хлорных каналах и т.п.
2. По строению (родству их химического строения и происхождения образующих их белков). По строению (структуре) и по происхождению от однотипных генов различные ИК объединяются в отдельные семейства. Например, выделяют три семейства лиганд-активируемых ИК: 1) семейство с пуриновыми рецепторами (АТФ-активируемые), 2) с никотиновыми АХ-рецепторами, ГАМК-, глицин- и серотонин-рецепторами, 3) с глутаматными рецепторами. При этом в одно и то же семейство попадают ИК с разной ионной селективностью, а также ИК с разными управляющими лигандами. Но зато образующие эти каналы белки имеют большое сходство в строении и происхождении.
3. По способу управления их состоянием. В этом случае мы будем говорить о потенциал-управляемых каналах, хемо-управляемых и т.д.
4. По связывающимся с ними лигандам (в том числе веществам-маркёрам) и т.д.
Создание удобной классификации является пока ещё не решённой проблемой. Как указывают Н.Н. Мушкамбаров и С.Л. Кузнецов, (2003), "в отличие от липидов, мембранные белки трудно классифицировать по их структуре. Более перспективно попытаться подразделить эти белки по их функциональной роли. Но и здесь нет законченной системы, т.к. любые попытки её создания наталкиваются на типичные трудности, когда один и тот же белок может быть отнесён к разным группам". Тем не менее, на наш взгляд, функциональная классификация ИК просто необходима для обучения студентов: биологов, медиков, психологов.
В основу предложенной нами функциональной классификации ИК (Сазонов В.Ф., 2011) положен способ управления их деятельностью, а не их селективная проницаемость к определённым ионам или химическое родство образующих их белков-каналоформеров. С этой точки зрения ИК делятся на неуправляемые и управляемые, т.е. либо постоянно открытые, либо открывающиеся-закрывающиеся при определённих воздействиях. Заметим, что большинство ИК являются управляемыми, но различаются между собой по механизмам управления. Воздействие регуляторного (управляющего) фактора на управляемый ИК вызывает конформационные изменения каналообразующих белков, канал открывается и ионы проходят по градиенту концентрации. При этом сам транспорт ионов через такие каналы не приводит к конформационным изменениям канальных белков и зависит только от разности концентраций веществ по обе стороны мембраны.
В одну и ту же функциональную группу нашей классификации могут попасть каналы различного молекулярного строения и с различной селективностью, т.е. пропускающие различные ионы. С другой стороны, сходные по строению и происхождению каналы могут оказаться в разных функциональных группах. Так, например, хлор-селективные ИК могут управляться как лигандами (глицином, ГАМК) и состоять в группе лиганд-управляемых каналов, так и потенциалом мембрнаны и состоять в группе потенциал-управляемых каналов (потенциал-активируемые хлорные каналы ClC).
Студентам
Для понимания электрических процессов, идущих в нервных клетках, формирования электрических потенциалов и нервных импульсов вполне достаточным будет разобраться в первых четырёх видах ионных каналов: 1) неуправляемыепостоянно пропускают через себя ионы калия, 2) потенциал-управляемые открываются при деполяризации и начинают в этих условиях пропускать через себя в клетку ионы натрия (в постсинаптических окончаниях и нервных отростках) или же ионы кальция (в пресинаптических окончаниях или рецепторных клетках), 3) хемо-управляемые открываются под действием медиатора и начинают пропускать через себя в клетку ионы натрия, что вызывает деполяризацию в виде возбуждающего постсинаптического потенциала (ВПСП), 4) стимул-управляемые находятся в сенсорных рецепторах (рецепторных клетках или рецепторных нервных окончаниях) и открываются под действием стимула (раздражителя), начиная пропускать через себя ионы натрия, что вызывает деполяризацию в виде рецепторного потенциала.
"Управление ионными каналами" - означает их открытие или закрытие под определёнными воздействиями.
Виды ионных каналов согласно функциональной классификации:
1. Неуправляемые (независимые, "проточные"). Конечно, это название условно и отражает лишь основное функциональное состояние подобных каналов. Пожалуй, полностью независимых и неуправляемых ИК в мембране просто не существует, и все они так или иначе регулируются. Неуправляемые ИК обычно находятся в постоянно открытом состоянии и обеспечивают постоянный ионный ток через открытую пору канала как в клетку, так и из клетки. Процесс перемещения ионов через такие ИК идёт пассивно за счёт диффузии под действием химических сил (по градиенту их концентрации) и/или электрических сил (по электрическому градиенту зарядов между внутренней и наружной сторонами мембраны).
Если неуправляемые каналы различают вещества только по размеру и пропускают через себя по градиенту концентрации все молекулы меньше определённой величины, т.е. служат фильтрами молекулярных размеров, то их называют "неселективные каналы", или "поры". Селективные каналы,работают избирательно и обеспечивают перенос только определённых ионов. Ионная селективность (избирательность) каналов определяется их диаметром и строением внутренней поверхности канала. Например, катионселективные каналы пропускают только катионы, так как содержат много отрицательно заряженных аминокислотных остатков.
Примеры: калиевые каналы утечки (они участвуют в формировании нервными клетками мембранного потенциала покоя), эпителиальные натриевые каналы ENaCs (они обеспечивают обратное всасывание ионов натрия в почках, прямой кишке, лёгких, потовых железах и пр., также обеспечивают восприятии солёного вкуса вкусовыми рецепторами во рту), анионные каналы эритроцитов (они пропускают анионы Сl-, OH-, HCO3-).
Видео: Калиевый ионный канал
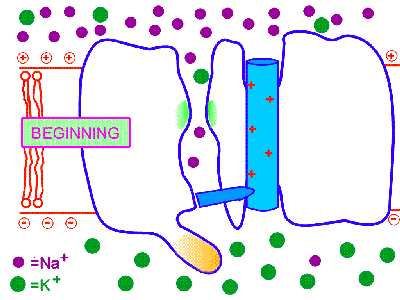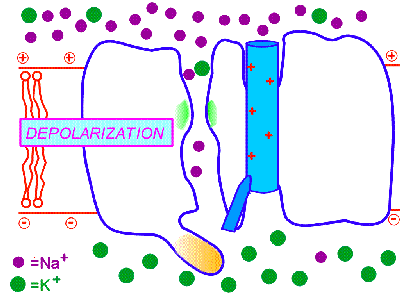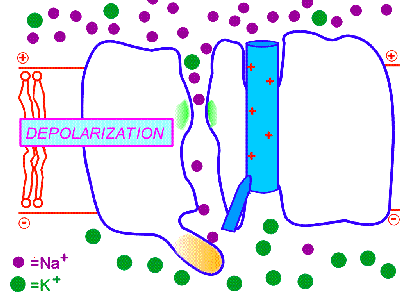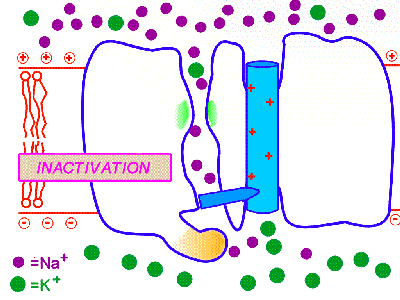
2. Потенциал-управляемые (потенциал-чувствительные, потенциал-зависимые, потенциал-активируемые, voltage-gated). Так, потенциал-управляемые натриевые каналы открываются под действием сдвига электрического потенциала мембраны, превышающего критический уровень деполяризации. Поэтому при достижении определённого порогового уровня деполяризации мембраны они открываются, а при обратном снижении уровня деполяризации - оказываются закрытыми. Но важно знать то, что ещё до обратного снижения уровня деполяризации эти каналы закрываются с внутренней стороны специальными белковыми "пробками" и это происходит автоматически, незасисимо от изменений деполяризации. Вследствие этого потенциал-управляемые натриевые ИК находятся в открытом состоянии всего несколько миллисекунд, а потом закрываются "пробкой", т.е. инактивируются. Окончательно они переходят в закрытое состояние при реполяризации и восстановлении потенциала покоя. Как при химической, так и при фармакологической модификации таких ИК у них сохраняется основной механизм активации и инактивации в ответ на сдвиг мембранного потенциала, что и определяет быстрые изменения катионной проницаемости возбудимых мембран за счёт потенциал-управляемых ИК. Именно такого типа потенциал-управляемые натриевые ИК обеспечивают перемещение нервного импульса по мембране нейрона (смотри: потенциал действия и нервный импульс). Такие потенциал-зависимые натриевые каналы I открываются на уровне КУД, т.е. -55mV, они и формируют потенциал действия и нервный импульс.
На рисунке справа - условная схема работы потенциал-управляемого ИК (кликните на рисунок, чтобы увидеть процесс в динамике).
3. Хемо-управляемые (хемочувствительные, хемозависимые, лиганд-управляемые, лиганд-зависимые, рецептор-активируемые). Они открываются при связывании с рецепторным участком канала специфического лиганда (управляющего вещества: трансмиттера или его миметика). Такие каналы обычно локализованы в химических синапсах на их постсинаптических мембранах и преобразуют химический сигнал, возникающий за счёт пресинаптического высвобождения нейромедиатора, в постсинаптический электрический локальный потенциал. Смотри: локальный потенциал, 3_3 Синапсы, синапсы, медиаторы и модуляторы.
Важно принять во внимание то, что хемо-управляемые (лиганд-управляемые) ионные каналы в качестве синонима называют также рецепторами.
Поэтому строение таких ионных каналов часто рассматривают там же, где рассматривается строение мембранных рецепторов.
Примеры: каналы с никотиновыми ацетилхолиновыми рецепторами nAChR), серотониновыми рецепторами (5-HT3), глициновыми, ГАМК-рецепторами (GABAA и GABAC).
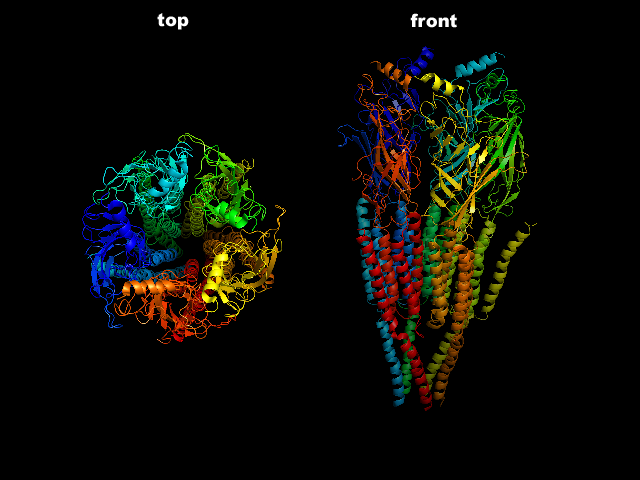
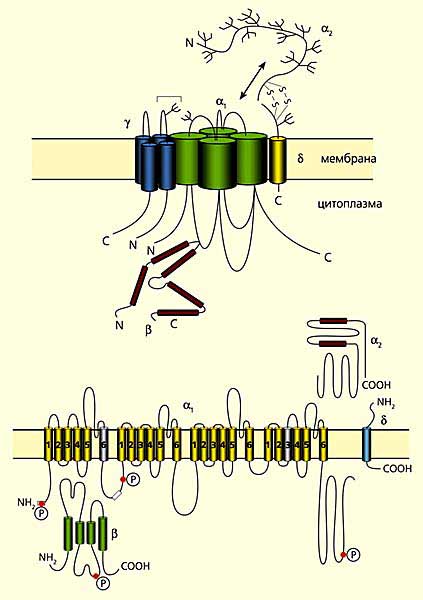
На рисунке справа - лиганд-управляемый ионный канал с никотиновым ацетилхолиновым рецептором (никотин - миметик, ацетилхолин - трансмиттер). Канал состоит из 5 субъединиц и поры в центре. Вверху — поперечный разрез этих субъединиц: a1, a2, b, g, d. Внизу — участки субъединиц, образующих «воротную систему» канала. Представлены аминокислотные последовательности М2 a-спирали в b- и d-. Из 5 субъединиц, образующих пору, изображены лишь 4, а ближайшая к нам удалена, чтобы были видны участки М2, облицовывающие ионный канал, и ворота. Видно, что большая часть молекулы белка выходит за пределы внешней поверхности плазматической мембраны, образуя молекулярные рецепторы к лиганду. Каждая из двух a-субъединица содержит связывающий центр для ацетилхолина, следовательно, с рецептором может связаться 2 молекулы лиганда (нейротрансмиттера или нейромиметика). Ворота, находящиеся в пределах поры, открываются при связывании ацетилхолина с рецепторным участком канала. Отрицательно заряженные остатки глутаминовой и аспарагиновой аминокислот (они выделены синим цветом) имеются в обоих концах М2-спиралей, т.е. с двух сторон поры, благодаря чему предотвращается вход в канал анионов, а катионы Na+ и К+ при его закрытом состоянии могут быть связаны уже в самом канале.
Видео: Работа хемо-управляемого (лиганд-управляемого) ионного канала
Они открываются под воздействием специфичного и адекватного для них стимула (раздражителя). Такие каналы обеспечивают сенсорное восприятие и располагаются в мембране сенсорных рецепторов.
В настоящее время стимул-управляемые механочувствительные ИК обнаружены не только в специализированных механорецепторных структурах, но также и в мембранах бактерий, грибов, растений, позвоночных и беспозвоночных животных. Механочувствительные каналы не только обеспечивают сенсорное восприятие механического раздражения, но также вовлечены в контроль клеточного цикла, регуляцию объёма и роста клеток, секрецию и эндоцитоз.
TRP-каналы в мембране терморецепторов кожи обеспечивают термотрансдукцию, открываясь при различных значениях темпераруры. Они пропускают катионы, особенно ионы кальция.
5. Совместно-управляемые (NMDA-рецепторно-канальный комплекс). Они открываются одновременно как лигандами, так и определённым электрическим потенциалом мембраны. Можно сказать, что у них двойное управление.
Пример: NMDA-рецепторно-канальный комплекс, имеющий сложную систему управления, включающую в себя 8 рецепторных участков-сайтов, с которыми могут связываться различные лиганды.
6.Опосредованно-управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер-управляемые, управляемые метаботропными рецепторами). Они открываются и закрываются не под действием прямых внешних сигналов, а вследствие опосредованного воздействия на нихвнутриклеточных вторичных мессенджеров (ионов кальция Са2+, цАМФ, цГМФ, ИФ3, диацилглицерола). Основной механизм такого управления - фосфорилирование ионного канала с внутренней стороны мембраны.
Опосредованное вторичными мессенджерами управление является не прямым, а вторичным. Оно зависит не только от внешнего воздействующего сигнала, но и от наличия, концентрации и активности вторичных мессенджеров. Пусковым сигналом к началу этого процесса может служить воздействие на так называемый метаботропный рецептор, не относящийся к структуре самого управляемого ионного канала и расположенный на мембране где-то отдельно от него. Воздействие на метаботропный рецептор приводит к повышению в клетке концентрации вторичных посредников-мессенджеров. Это ионы Са2+, цАМФ, цГМФ, ИФ3, диацилглицерол. Они активируют соответствующие ферменты-протеинкиназы: А-киназы (цАМФ-зависимые), G-киназы (цГМФ-зависимые), В-киназы (кальций-кальмодулин-зависимые) или С-киназы (кальций-фосфолипид-зависимые). В свою очередь, активированные киназы фосфорилируют ИК мембраны изнутри клетки, т.е. присоединяют к ним фосфаты. В результате этого канал может перейди надолго в новое состояние (открытое или, наоборот, закрытое). После срезания фосфатов ферментом фосфатазой канал возвращается к своему прежнему состоянию. В некоторых случаях такой вторичный мессенджер, как G-белок в виде своей активной субъединицы бета-гамма может сам присоединиться к ионному каналу и поменять его состояние. Так, например, могут открываться (активироваться) калиевые каналы при раздражении ацетилхолином мускариновых рецепторов, связанных с G-белком.
Вот, например, опосредованные ион-управляемые кальций-активируемые хлорные каналы являются одним из основных компонентов системы эпителиальной секреции, сенсорной трансдукции , регулирования нейронной и сердечной возбудимости у животных. В клетках растений кальций-активируемые хлорные каналы ответственны за состояние тургора клетки. При повышении концентрации ионов кальция внутри клетки эти каналы открываются и начинают пропускать ионы хлора.
К опосредованно-управляемым (мессенджер-управляемым) ионным каналам формально можно отнести также все каналы, которые управляются "изнутри" с помощью посредников - вторичных мессенджеров. Такой способ управления чаще всего является дополнительным по отношению к "внешнему управлению", и получается, что к мессенджер-управляемым каналам относится большинство каналов из других групп нашей функциональной классификации. Например, все те ИК, которые могут подвергаться фосфорилированию.
7. Актин-управляемые (актин-регулируемые, actin-regulated, actin-gated channels). Они открываются и закрываются за счёт разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков.
В электроневозбудимых клетках активация и инактивация актин-управляемых потенциал-независимых натриевых каналов контролируется процессами разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков. Актиновые элементы цитоскелета, по-видимому, представляют важнейшую часть потенциал-независимого воротного механизма, управляющего открыванием и закрыванием каналов. Именно сборка микрофиламентов на цитоплазматической стороне мембраны приводит к инактивации таких каналов.
8. Коннексоны (двойные поры). Образуют в мембранах контактирующих клеток сквозные непрерывные каналы через две мембраны сразу в зоне щелевых контактов для взаимного обмена веществами между этими клетками. Через коннексоны передаются электрические сигналы, аминокислоты и небольшие молекулы управляющих веществ: цАМФ, InsP3, аденозин, АДФ и АТФ. Они состоят из 6 белковых субъединиц (коннексинов), живущих всего несколько часов. Коннексины - это политопные интегральные мембранные белки 4 раза прошивающие мембрану, имеющие две внеклеточные петли (EL-1 и EL-2), цитоплазматическую петлю (CL) с N-концом (AT) и C-концом (CT), вдающимися в цитоплазму. Через коннексоны соединяется внутренняя среда соседствующих клеток.
Коннексоны являются "неспецифически-управляемыми" каналами. Их состояние регулируется pH, электрическим потенциалом, ионами Са2+, фосфорилированием и другими факторами.
Коннексоны найдены практически во всех видах клеток.
9. «Энерго-зависимые транспортёры» (ионные насосы, ионные помпы, ионные обменники, транспортёры). Это особая группа динамичных пор, проводящих ионы через мембрану, которые формально не относятся к ИК. Их деятельность обеспечивается энергией расщепления АТФ. Они представлены мембранными ферментными белками АТФазами, которые активно протаскивают через себя ионы, используя для этого энергию расщепления АТФ, и обеспечивают активный транспорт ионов через мембрану даже против их градиента концентрации.
Глутаматные лиганд-управляемые (хемозависимые) и совместно-управляемые ионные каналы
Постсинаптические рецепторы к глутамату классифицируются в соответствии с аффинностью (сродством) к трем экзогенным агонистам:
1) AMPA-рецепторы, активируются α-аминометилизоксазолилпропионовой кислотой, а также квисгулатом.
2) KA-рецепторы, активируются каиновой кислотой.
3) NMDA-рецепторы, активируются N-метил-D-аспартатом. Подробнее: Перейти
Ионные каналы, активируемые квисгулатом и каинатом, подобны каналам, которые управляются никотиновыми рецепторами — они пропускают смесь катионов (Na+ и К+). По нашей функциональной классификации они являются лиганд-управляемыми.
Стимуляция NMDA-рецепторов имеет сложный характер активации: ионный ток, который переносится не только Na+ и К+,но также Са++ при открывании ионного канала рецептора, зависит от потенциала мембраны. По нашей классификации они относятся к совместно-управляемым ионным каналам.
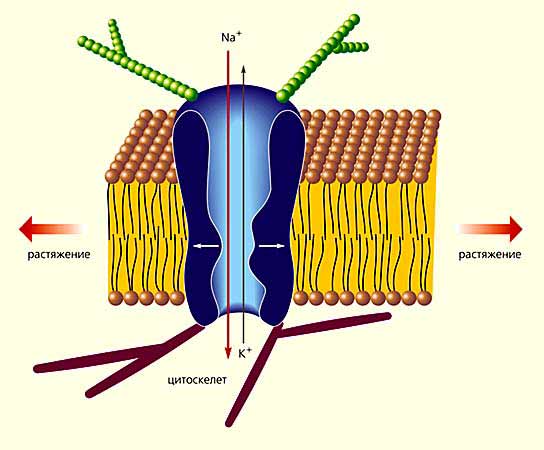
Потенциал-управляемые ионные каналы
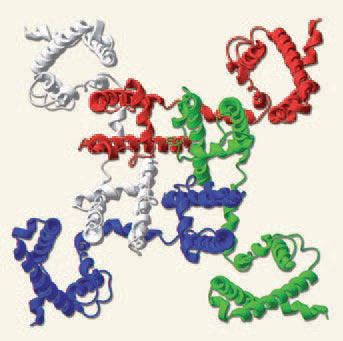
На рисунке слева представлена модель, отражающая взаимодействие субъединиц потенциал-управляемого кальциевого канала (вверху), и его доменная структура.
Этот белковый канал построен из пяти субъединиц: a1, a2, b, g и d. Три из них (a1, g и d) прошивают плазматическую мембрану, причем их N- и C-концы расположены в цитоплазме. b-субъединица полностью находится в цитозольном пространстве клетки, а a2-субъединица, взаимодействующая через S-S-мостики с d-субъединицей, выступает над наружной стороной мембраны. Все субъединицы, за исключением d, состоят из нескольких компактных доменов, соединенных между собой петлями. В виде цилиндров изображены a-спиральные участки субъединиц. Места фосфорилирования (Р) аминокислотных остатков показаны красным цветом.
(Источник: Фундаментальная и клиническая физиология. Под ред. А.Г. Камкина и А.А. Каменского. М.: Академия, 2004. 1072 с.)
Кальциевые ионные каналы
Особое значение имеют для нейронов кальциевые ИК, потому что кальций - это не только положительно заряженный ион, но и мощный биологический внутриклеточный активатор. Попадая в нейрон через открывшиеся для него каналы, кальций запускает каскад биохимических реакций в нейроне, приводящих к самым разным результатам. Результаты зависят от того, какие именно белки встретятся с кальцием внутри клетки.
Порог раскрытия для кальциевых каналов Т-типа у разных нейронов ЦНС лежит в диапазоне от -83 до -50 мВ [Perez-Reyes E. (2003) Molecular physiology of low-voltage-activated t-type calcium channels. Physiol. Rev., 83(1):117-61. Gutnick, MJ; Yarom, Y (1989) Low threshold calcium spikes, intrinsic neuronal oscillation and rhythm generation in the CNS. Journal of Neuroscience Methods. 28 (1–2): 93–99. Huguenard JR (1996) Low-threshold calcium currents in central nervous system neurons. Annual review of physiology 58 (1), 329-348. Tsien RW, Lipscombe D, Madison DV, Bley KR, Fox AP. (1988) Multiple types of neuronal calcium channels and their selective modulation. Trends Neurosci. 11(10):431-8.].
Это означает, что нейроны могут активироваться входящими ионами кальция на подпороговом уровне, ещё до генерации нервного импульса.
Ионные каналы мембраны и их виды
Ионные каналы (ИК) относятся к мембранным интегральным белкам, пронизывающим клеточную мембрану поперёк. Они выстроены из нескольких субъединиц и образуют структуру со сложной пространственной конфигурацией. В этой своеобразной "молекулярной машине" имеются системы открытия, закрытия, избирательности, активации, инактивации и регуляции, а также участки-сайты для связывания с управляющими веществами. В ИК (например, в NMDA-комплексе) может встречаться до 8 точек для приложения управляющего воздействия, т.е. для связывания управляющих веществ, меняющих состояние канала. Отдельные части этой структуры являются подвижными относительно друг друга, и они непрерывно шевелятся, меняя своё взаиморасположение. Поэтому общее состояние ИК может непрерывно меняться, и он в определённые моменты времени может оказаться в открытом состоянии, а в другие моменты — в закрытом. Это вероятностный процесс, и внешнее управление каналом лишь меняет вероятность его нахождения в открытом и закрытом состоянии. Для ИК функционально важными являются именно эти их переходы между открытыми и закрытыми состояниями. Эти переходы совершаются практически моментально. Иногда канал открыт только одну миллисекунду или даже меньше, хотя в следующий раз он может открыться на гораздо более продолжительное время. Тем не менее, каждый канал имеет своё характерное среднее время открытого состояния, и все вариации происходят именно вокруг этого среднего показателя. Некоторые ИК открываются достаточно часто даже в покое. Это означает, что вероятность нахождения таких каналов в открытом состоянии даже в неактивированной клетке относительно высока. Большинство таких постоянно открытых ионных каналов проницаемо для калия или хлора. Они важны для генерации мембранного потенциала покоя. Такие ИК мы предлагаем называть «неуправляемыми». Другие ИК мембраны, которых большинство, большую часть времени закрыты, то есть вероятность нахождения их в открытом состоянии очень низка. Активация этих каналов адекватным стимулом резко увеличивает вероятность их открытия, хотя и нельзя сказать, что эта вероятность достигает 100%, и что каждый активируемый канал обязательно откроется. Этот же стимул может деактивировать ИК, бывшие активными в покое. Важно помнить, что активация или деактивация канала означает возрастание или снижение вероятности открытия канала, но не увеличение или уменьшение продолжительности времени открытого состояния канала. Такие ИК мы предлагаем называть в целом «управляемыми». Помимо активации и деактивации, прохождение ионов через каналы регулируется двумя другими факторами, которые можно назвать «инактивацией» и «блокированием». Инактивация заключается в том, что ионный канал переходит в новое конформационное состояние, в котором обычный активирующий стимул не способен вызвать открытие канала. Для ионных каналов, активируемых деполяризацией, такое состояние называется инактивацией. Для каналов, отвечающих на химические стимулы, это состояние известно как «десенситизация», т.е. понижение чувствительности. Второй механизм - блокирование открытого канала. Такое случается, когда, например, крупная молекула (например, токсин) связывается с ионным каналом и физически закупоривает пору. Другим примером может служить блокирование некоторых катионных каналов ионами магния Mg2+. В этом случае ионы магния сами не проходят через ионный канал на другую сторону мембраны, но связываются с каналом в области его устья и тем самым мешают прохождению через него других катионов. Такое явление называется «магниевая пробка». Некоторые каналы специфически отвечают на изменения в состоянии клеточной мембраны нейрона. Наиболее яркими представителями этой группы являются потенциал-активируемые каналы, реагирующие на электрический заряд мембраны. Примером может служить чувствительный к потенциалу натриевый канал, который отвечает за самоускоряющуюся деполяризацию, лежащую в основе развития потенциала действия. Такие ИК мы предлагаем называть «потенциал-управляемыми». К группе управляемых каналов относятся также механочувствительные ИК, которые повышают свою проницаемость в ответ на механические воздействия на клеточную мембрану. Сенсорные рецепторы растяжения, содержащие в своей мембране такие ИК, найдены, например, в механорецепторах кожи. Такие ИК мы предлагаем называть «стимул-управляемыми». Другие ИК открываются тогда, когда определённые химические вещества (их обобщённое название - «лиганды») активируют связывающие центры на молекуле канала. Такие лиганд-активируемые ИК подразделяются на две подгруппы, в зависимости от того, являются ли активные центры внутриклеточными или внеклеточными. Каналом, отвечающим на внеклеточную активацию, является, например, катионный канал постсинаптической мембраны в скелетной мышце. Этот канал активируется нейротрансмиттером ацетилхолином, освобождающимся из двигательного нервного окончания. Открытие ацетилхолин-активируемого ионного канала позволяет ионам натрия войти в клетку, вызывая деполяризацию мышечного волокна. Такие ИК мы предлагаем называть «лиганд-управляемыми». Лиганд-активируемые каналы, отвечающие на внутриклеточные стимулы, включают в себя каналы, чувствительные к местным изменениям концентрации специфических ионов. Например, кальций-активируемые калиевые каналы активируются локальным повышением концентрации внутриклеточного кальция. Такие каналы играют важную роль в реполяризации клеточной мембраны во время завершения потенциала действия. Помимо ионов кальция, типичными представителями лигандов, активирующих ИК с цитоплазматической стороны мембраны, являются циклические нуклеотиды. Циклический ГМФ, например, отвечает за активацию натриевых каналов в палочках сетчатки. Такой тип канала играет принципиальную роль в работе зрительного анализатора. Такие ИК мы предлагаем называть «опосредованно-управляемыми» ( «мессенджер-управляемыми»). Предлагаемая нами классификация ИК по способу управления их состоянием не является достаточно строгой, посокльку выделяемые нами группы не являются взаимоисключающими, и каналы могут одновременно входить в несколько групп. Например, кальций-активируемые калиевые каналы (опосредованно-управляемые) чувствительны также и к изменению потенциала (т.е. являются одновременно и потенциал-управляемыми или же совместно-управляемыми), а некоторые потенциал-управляемые ИК чувствительны к внутриклеточным лигандам, т.е являются дополнительно опосредованно-управляемыми. Что тут поделаешь? Реальность часто бывает сложнее, чем наши теории на её счёт! Следует также помнить, что ИК различаются по своей избирательности (селективности). Так, некоторые катионные каналы пропускают в основном только один вид ионов: натрий, калий или кальций, другие же являются менее избирательными. Анионные каналы сравнительно не избирательны для малых анионов, но они пропускают в основном ионы хлора, так как хлор является самым распространенным анионом в составе внеклеточной и внутриклеточной жидкости в организме.
Используемая литература:
1. Ионные каналы возбудимой клетки (структура, функция, патология) / Зефиров А.Л., Ситдикова Г.Ф. Казань: Арт-кафе, 2010. 271 с.
2. Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. Учебное пособие для студентов медицинских вузов. М.: ООО "Медицинское информационное агентство", 2003. 544 с.
3. Сазонов В.Ф. Функциональная классификация мембранных ионных каналов // Научные труды III Съезда физиологов СНГ / Под ред. А.И. Григорьева, О.А. Крышталя, Ю.В. Наточина, Р.И. Сепиашвили. М.: Медицина–Здоровье, 2011. С. 72. (Электронная версия: physiology-cis.org/Page181.html)
4. Фундаментальная и клиническая физиология. Под ред. А.Г. Камкина и А.А. Каменского. М.: Академия, 2004. 1072 с.
На сайте введена регистрация через социальные сети, если вы хотите оставлять комментарии без потверждения, пожалуйста, воспользуйтесь именно этим типом аутентификации.
Если у вас уже есть аккаунт на сайте, вы можете привязать его к любой социальной сети? зайдя в настройки вашего аккаунта("Мои учётные данные") ниже и воспользовавшись вкладкой "Подключение к социальным сетям".
После того, как вы зайдёте при помощи аккаунта в социальной сети, ваши возможности на сайте возрастут.
Поддержка сайта
Вы можете поддержать сайт не только добрым словом, но и материально!
Это очень поможет. IT-специалисты, следящие за сайтом день и ночь, хотя бы лишнюю чашечку кофе выпьют.
Для этого по своему желанию перечислите любую сумму на карту Сбербанка номер:










 ВКонтакте
ВКонтакте Facebook
Facebook Twitter
Twitter{kind=link}
{kind=link}